
Evolución de Biston betularia
- Este artigo trata sobre a importancia da Biston betularia na bioloxía evolutiva. Para a evolución a partir dos seus antepasados ver Evolución dos insectos


A evolución de Biston betularia é un exemplo evolutivo de cambio direccional de cor nunha poboación de avelaíñas como consecuencia da contaminación do aire durante a Revolución Industrial. A frecuencia das avelaíñas desta especie de cor escura incrementouse en dita época histórica, un exemplo do que se denominou melanismo industrial. Posteriormente, cando a contaminación se reduciu, a forma de cor clara pasou a predominar outra vez. O melanismo industrial na Biston betularia foi unha das primeiras comprobacións da selección natural de Charles Darwin en acción, e converteuse nun exemplo clásico na ensinanza da evolución.[1][2] En 1978 Sewall Wright describiuno como "o caso máis claro no cal un proceso evolutivo notorio foi observado realmente."[3][4]
A forma de cor escura ou melánica da B. betularia (var. carbonaria) non se coñecía antes de 1811. Despois dunha recolección no campo en 1848 en Manchester, unha cidade industrial de Inglaterra, observouse que esa variedade se incrementara drasticamente. Ao final do século XIX xa superaba á variedade orixinal de tipo claro (var. typica), chegando ao 98% en 1895.[5] Da importancia evolutiva desta avelaíña só se fixeron especulacións durante a vida de Darwin. Pero 14 anos despois da vida de Darwin, en 1896, J.W. Tutt presentouna como un caso de selección natural.[6] Debido a isto, a idea espallouse grandemente, e máis persoas quedaron convencidas da teoría de Darwin.
Bernard Kettlewell foi o primeiro que investigou o mecanismo evolutivo que estaba detrás da adaptación da B. betularia, entre 1953 e 1956. Atopou que un corpo e ás de cores claras era unha camuflaxe efectiva nun ambiente limpo, como en Dorset, mentres que as cores escuras eran beneficiosas nun ambiente contaminado como Birmingham. Esta supervivencia selectiva era debida a que os predadores das avelaíñas (aves) capturaban con maior facilidade as avelaíñas escuras sobre as árbores claras (non lixadas pola contaminación do feluxe). Este caso, apoiado no experimento de Kettlewell, converteuse nun exemplo canónico da evolución darwinista e evidencia da selección natural usada nos libros de texto estándar.[7]
Porén, como Theodore David Sargent non conseguiu replicar o experimento e criticou os métodos de Kettlewell a finais da década de 1960, creceu o escepticismo sobre o caso. Cando se publicou o libro de Judith Hooper Of Moths and Men (De avelaíñas e homes) en 2002, o experimento de Kettlewell foi atacado máis duramente, acusado de fraude e suscitou un amplo rexeitamento. Estas críticas foron aproveitadas como un importante argumento polos creacionistas, que negan a realidade da evolución. Michael Majerus foi o principal defensor do exemplo ao realizar un amplo experimento que durou sete anos. O experimento de Majerus comezou en 2001, é o máis elaborado do seu tipo en bioloxía de poboacións, e os seus resultados foron publicados postumamente en 2012, e vindicaban o traballo de Kettlewell en gran detalle. Isto restaurou a validez do exemplo da evolución da B. betularia como "a evidencia máis directa" e "un dos exemplos máis claros e doados de entender de evolución darwinista en acción".[8]
Orixe e evolución
[editar | editar a fonte]
Antes da Revolución Industrial, a B. betularia escura era rara. Os primeiros espécimes negros (de orixe descoñecida) foron gardados na Universidade de Oxford en 1811.[9][10][11] O primeiro espécime vivo capturouno R.S. Edleston en Manchester, Inglaterra en 1848, pero informou non disto ata 16 anos despois en 1864 na revista Entomologist.[12] Edleston indicou que en 1864 era o tipo máis común de avelaíña no seu xardín de Manchester. As avelaíñas claras podían confundirse cos liques de cores claras e a codia das árbores, e a avelaíña inicialmente menos común negra tiña máis probabilidades de ser detectada e comida polas aves. Como resultado de que os liques e as árbores inglesas eran claras, as avelaíñas de cor clara eran moito máis efectivas á hora de ocultarse ante os depredadores e a frecuencia do alelo escuro era ao principio só do 0,01%.[13]
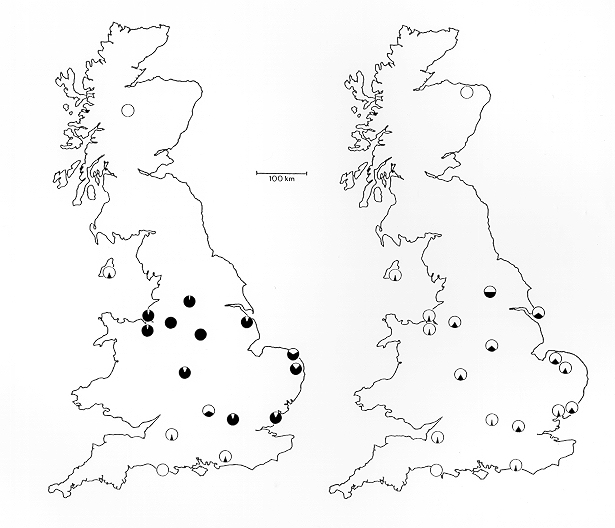
Durante as primeiras décadas da Revolución Industrial en Inglaterra, o campo entre Londres e Manchester quedou cuberto de feluxe procedente das novas fábricas que queimaban carbón. Moitos dos liques claros morreron polas emisións de dióxido de xofre e as árbores quedaron escurecidas. Isto causou un incremento da predación polas aves sobre as avelaíñas de cores claras, que xa non se confundían tan ben no seu ecosistema contaminado: de feito, os seus corpos contrastaban agora fortemente coa cor escurecida da codia das árbores. As avelaíñas de cores escuras camuflábanse agora moi ben co novo estado das árbores.[7] A poboación de avelaíñas de cores escuras incrementáronse rapidamente. A metade do século XIX, o número de avelaíñas de cores escuras aumentara notablemente, e en 1895, a porcentaxe de avelaíñas de cores escuras en Manchester era do 98%, un forte cambio (de case o 100%) respecto da frecuencia orixinal.[7] Este efecto da industrialización na cor corporal levou a que se acuñase o termo "melanismo industrial".[2]
Xa durante a vida de Darwin se percibira que o melanismo industrial podía ser unha evidencia que apoiase a teoría da selección natural de Charles Darwin. Albert Brydges Farn (1841–1921), un entomólogo británico, escribiulle a Darwin o 18 de novembro de 1878 para discutir a súa observación das variacións de cor na avelaíña Gnophos obscurata (agora chamada Charissa obscurata). Decatouse da existencia de avelaíñas escuras na turba do New Forest, avelaíñas marróns sobre solos de arxila e solos vermellos de Herefordshire e brancas sobre os cantís de creta de Lewes; entón suxeriu que esta variación era un exemplo de "supervivencia dos máis adaptados". Díxolle a Darwin que atopara avelaíñas escuras nas ladeiras de creta onde a follaxe quedara escurecida polo fume procedente de fornos de cal, e tamén oíra que as avelaíñas brancas se fixeran menos comúns en Lewes despois de que empezasen a funcionar os fornos de cal había algúns anos.[14] Darwin parece que non respondeu a esta información, posiblemente porque pensaba que a selección natural tiña que ser un proceso moito máis lento.[15] Non se publicou unha explicación científica da coloración das avelaíñas ata 1896, 14 anos despois da morte de Darwin, cando J.W. Tutt relacionou explicitamente o melanismo de B. betularia coa selección natural.[13]
Aumento e diminución da frecuencia dos fenotipos
[editar | editar a fonte]O melanismo aparecera nas poboacións de Europa e Norteamérica de B. betularia. A información sobre o aumento de frecuencia é escasa. Sábese moito máis sobre a posterior caída na frecuencia dese fenotipo, xa que isto foi medido polos estudosos dos lepidópteros utilizando trampas para avelaíñas.
Steward compilou datos dos primeiros rexistros da B. betularia por localidade e deduciu que o morfo carbonaria era o resultado dunha soa mutación que se espallou posteriormente. En 1895, alcanzou unha frecuencia do 98% en Manchester.[16]
Desde aproximadamente 1962 ata o presente, a frecuencia do fenotipo carbonaria estivo baixando de forma sostida en liña coa maior limpeza do aire arredor das cidades industriais. O seu declive foi medido con máis precisión que o seu aumento, por medio de estudos científicos máis rigorosos. Especialmente, Bernard Kettlewell realizou unha avaliación nacional en 1956, Bruce Grant realizou outra similar a inicios de 1996,[17] e L.M. Cook outra en 2003.[18]
Resultados similares atopáronse en América. As formas melánicas non se atoparon no Xapón. Crese que isto se debe a que as B. betularia no Xapón non habitan en rexións industrializadas.[19]
Xenética
[editar | editar a fonte]J.W. Tutt foi o primeiro que propuxo a "hipótese de predación diferencial polas aves" en 1896, como mecanismo de selección natural. Os morfos melánicos estaban mellor camuflados sobre a codia das árbores sen liques foliosos, mentres que os morfos typica estaban mellor camuflados contra árbores con liques. Como resultado, as aves atoparían e comerían máis frecuentemente os morfos que non se camuflasen ben.[20]
En 1924, J.B.S. Haldane calculou, usando un modelo de selcción xeral simple, a vantaxe selectiva necesaria para a evolución natural rexistrada de B. betularia, baseándose na asunción de que en 1848 a frecuencia das avelaíñas escuras era do 2%, e en 1895 era do 95%. A forma escura ou melánica tería que estar un 50% mellor adaptada que a típica de cor clara. Mesmo considerando posibles erros no modelo, isto exclúe razoablemente o proceso estocástico da deriva xenética, porque os cambios eran demasiado rápidos.[21] A análise estatística de Haldane da selección natural da variante melánica en B. betularia converteuse nunha parte ben coñecida dos seus traballos para demostrar os modelos matemáticos que combinaba a selección natural coa xenética mendeliana podía explicar a evolución, un esforzo que xogou un papel clave na fundación da disciplina da xenética de poboacións e o principio da síntese moderna da teoría evolutiva coa xenética.[22]
En B. betularia o alelo para ter cor escura é dominante, mentres que o alelo das avelaíñas de corpo claro é recesivo, o que significa que as avelaíñas typica teñen un fenotipo (características visible ou detectables) que só se ve no xenotipo homocigoto (un organismo que ten dúas copias do mesmo alelo), e nunca no heterocigoto. Isto serve para explicar a forma tan rápida coa que a poboación cambia cando está sendo seleccionada pola coloración escura.
A Biston betularia é tamén un modelo de evolución paralela na incidencia do melanismo na forma británica (f. carbonaria) e a forma americana (f. swettaria), xa que son indistinguibles en aparencia. A análise xenética indica que ambos os fenotipos se herdan como trazos autosómicos dominantes. As hibridacións cruzadas indican que os fenotipos se producen por isoalelos nun só locus.[23]
O xene para carbonaria en B. betularia pensábase que estaba nunha rexión do seu cromosoma 17, pero máis tarde chegouse á conclusión que non podía contelo porque ningún dos xenes do cromosoma codificaba nin o padrón das ás nin a melanización.[24][25] A rexión que foi utilizada para encontralo foi o primeiro intrón do ortólogo do xene cortex de Drosophila. Por medio de ir eliminando candidatos dentro desa rexión baseándose na súa rareza, quedou unha inserción de 21925 pares de bases.[25]
A inserción denominada carb-TE, é un elemento transpoñible de clase II que ten aproximadamente 9-kb de secuencia non repetitiva que está repetida en tándem dúas e tres veces.[25] Hai seis pares de bases de repeticións invertidas e 4 pares de bases duplicadas no sitio diana que non están presentes en avelaíñas typica. Carb-TE ten unha expresión máis alta durante o estadio de rápida morfoxénese do disco alar.[25] O mecanismo de como incrementou a expresión ou se é o único xene implicado aínda non se coñece.[25]
Hipóteses alternativas
[editar | editar a fonte]Propuxéronse varias hipóteses alternativas á selección natural como forza impulsora da evolución durante as décadas de 1920 e 1930. A mutación aleatoria, a migración ou deriva xenética tamén foron consideradas forzas importantes da evolución.[26] Porén, a magnitude dos cambios observados só poden ser explicados pola selección natural. Pode verse en xenética de poboacións que un cambio non diferencial non causa evolución. Se as frecuencias alélicas son denominadas cos termos alxebraicos p e q, e dicimos que p = 0,6 e q = 0,4, entón unha redución non diferencial no tamaño da poboación de, digamos, 2000 a 100 individuos, aínda produce os mesmos valores de (aproximadamente) p = 0,6 e q = 0,4.
P.A. Riley propuxo un factor selectivo adicional, no que a quelación de metais pesados pola melanina pode protexer esta avelaíña contra os efectos tóxicos de metais pesados asociados coa industrialización. Esta vantaxe selectiva suplementaría o principal mecanismo selectivo de predación diferencial polas aves.[27]
Indución fenotípica
[editar | editar a fonte]John William Heslop-Harrison (1920) rexeitou a hipótese da depredación diferencial polas aves de Tutt baseándose en que non cría que as aves comesen as avelaíñas. En vez diso apoiaba a idea de que os contaminantes podían causar cambios no soma e plasma xerminal do organismo. Esta hipótese probablemente tiña as súas raíces na década de 1890, cando foi proposto como unha forma de lamarckismo. É importante sinalar o seu contexto histórico.[28]
Hasebroek (1925) foi o primeiro que puxo a proba esta hipótese. Sostiña que a contaminación do aire alteraba a fisioloxía dos lepìdópteros, producindo así un exceso de pigmento negro. Expuxo pupas de lepidópteros a varias doses de gases contaminantes, concretamente sulfuro de hidróxeno (H2S), amoníaco (NH3) e "piredina" (presumiblemente a forma en que ortografou piridina). Utilizou oito especies nos seus estudos, catro das cales eran especies de bolboretas que non mostraban melanismo. Ford (1964) afirmou que as ilustracións de Hasebroek mostraban que as formas anormais que aparecían non eran melánicas e Hasebroek non conseguiu estudar a súa xenética.[29][30]
Heslop-Harrison (Harrison e Garrett 1926; Harrison 1928) suxeriu que o incremento de avelaíñas melánicas en rexións industrializadas debíase á "presión de mutación", non á selección polos predadores, que se consideraba insignificante. Os sales de chumbo e manganeso estaban presentes nas partículas contaminantes transportadas polo aire e sueriu que estas causaban mutacións en xenes para a produción de melanina, pero non noutros. Utilizou como material as especies Selenia bilunaria e Tephrosia bistortata. As larvas foron alimentadas con follas que incorporaran eses sales e as melánicas aparecían seguidamente. Os experimentos similares de Hughes McKenney (1932) e Thomasen e Lemche (1933) fracasaron á hora de replicar estes resultados. Porén, o estatístico e xenetista Ronald Fisher mostrou que os controis de Heslop-Harrison eran inadecuados.[31] Esta hipótese, porén, parece que foi falsada por medio de experimentos de cruzamentos.
O experimento de Kettlewell
[editar | editar a fonte]- Artigo principal: Experimento de Kettlewell.
Os primeiros experimentos importantes sobre B. betularia foron levados a cabo por Bernard Kettlewell na Universidade de Oxford, baixo supervisión de E. B. Ford, que lle axudou a obter unha bolsa da Nuffield Foundation para realizar o experimento. En 1953, Kettlewell comezou un experimento preliminar no cal as avelaíñas eran liberadas nun gran aviario (de 18 m × 6 m), onde eran comidas polo ferreiriño Parus major. O seu principal experimento, feito na Reserva Natural de Cadbury en Birmingham, Inglaterra, consisitiu en marcar, liberar e recapturar as avelaíñas marcadas. Atopou que neste bosque contaminado as aveláiñas typica eran depredadas preferencialmente. Isto demostraba que o fenotipo melánico era importante para a supervivencia de B. betularia nese hábitat. Kettlewell repetiu o experimento en 1955 nun bosque non contaminado en Dorset e de novo nos bosques contaminados de Birmingham. Foi acompañado por Nico Tinbergen e fixeron unha película xuntos.[2][20] En 1956 repetiu os experimentos e atopou resultados similares; en Birmingham as aves comían a maioría das avelaíñas claras (75%), mentres que en Dorset comían as escuras (86%).[1][32]
Críticas
[editar | editar a fonte]Theodore David Sargent realizou experimentos entre 1965 e 1969, dos que sacou a conclusión de que non era posible reproducir os resultados de Kettlewell e dixo que as aves non mostraban preferencia sobre avelaíñas pousadas en troncos claros ou escuros.[33][34] Suxeriu que Kettlewell adestrara as aves para que capturasen as aves nos troncos para obter os resultados desexados.[3][35]
O libro de Michael Majerus de 1998 Melanism: Evolution in Action (Melanismo: Evolución en acción) é unha adaptación do libro de Kettlewell The Evolution of Melanism (A evolución do melanismo), no que se discuten as críticas dos métodos experimentais de Kettlewell.[20] Ao revisar este libro, Jerry Coyne concluíu que "polo momento debemos descartar a Biston como un exemplo ben coñecido de selección natural en acción, aínda que é un caso claro de evolución. Hai moitos estudos máis apropiados para o uso en clase."[36] Posteriormente, cando se publicaron os estudos completos de Marjerus en 2012, cambiou de opinión sobre a non idoneidade do caso da B. betularia como exemplo de evolución, aceptando plenamente o exemplo."[37]
O libro de Judith Hooper de 2002 Of Moths and Men (De avelaíñas e homes) é unha das críticas máis severas do experimento de Kettlewell.[38] Hooper argumentou que non se puideron encontrar as notas de campo de Kettlewell e suxeriu que o seu experimento era fraudulento, baseándose nas críticas de Sargent que alegaban que as fotografías de avelaíñas foron tomadas de avelaíñas mortas situadas nun tronco. Dixo que E. B. Ford era un "fanático darwinista",[39] e afirmou que se aproveitou do cientificamente inxenuo Kettlewell para obter os resultados experimentais desexados. Ela despois afirmou que os científicos en xeral mostraban unha aceptación "crédula e nesgada" da evolución.[40] A recepción do libro orixinou afirmacións de que a historia da evolución de B. betularia debería ser eliminada dos libros de texto.[41][42] Os científicos que examinaron as alegacións feitas por Hooper atoparon que carecían de mérito.[18][43][44]
.jpg)
Phillip E. Johnson, un cofundador do movemento creacionista do deseño intelixente, dixo que as avelaíñas "non se pousan nos troncos das árbores", "as avelaíñas eran pegadas aos troncos" para facer fotografías e que os experimentos eran "fraudulentos" e unha "estafa."[45] O partidario do deseño intelixente Jonathan Wells escribiu un ensaio sobre o asunto, unha versión acurtada do cal apareceu no número do 24 de maio de 1999 de The Scientist, afirmando que "en 25 anos de traballo de campo, C.A. Clarke e os seus colegas atoparon só unha B. betularia nun tronco de árbore", e concluíu que "o feito de que as B. betularia non descansan normalmente nos troncos de árbores invalida os experimentos de Kettlewell".[46] Wells despois escribiu en 2000 no libro Icons of Evolution (Iconas da evolución), no cal afirmaba, "porén, o que non explican os libros é que os biólogos coñecían desde a década de 1980 que a historia clásica tiña algúns serios defectos. O máis serio é que as B. betularia na natureza nin sequera descansan sobre os troncos das árbores. As fotografías de libro de texto resultou que foron escenificadas."[47] Porén, as B. betularia si descansan nos troncos en ocasións (experimentos posteriores indicaron que o facían nun 37% das veces) e as fotos obtidas en condicións naturais son practicamente iguais que as consideradas 'escenificadas'.[48]
Experimento de Majerus
[editar | editar a fonte]Desde 2001 a 2007, Majerus levou a cabo experimentos en Cambridge para resolver as varias críticas válidas do experimento de Kettlewell. Durante este experimento, observou as posicións naturais de descanso das B. betularia. De 135 avelaíñas examinadas aproximadamente a metade estaban nas pólas grandes das árbores, principalmente na metade inferior da póla, o 37% estaban sobre os troncos das árbores, principalmente no lado que dá ao norte, e só o 12,6% descansaban en ou baixo poliñas pequenas. En correspondencoa con Hooper engadiu un experimento para encontrar se os morcegos, e non as aves, podían ser os principais depredadores desas avelaíñas. Observou varias especies de aves que realmente depredaban as avelaíñas, e que a predación diferencial polas aves era un factor principal responsable do declive na frecuencia de carbonaria comparada con typica.[20] Describiu os seus resultados como unha reivindicación completa da teoría da selección natural da evolución da B. betularia e dixo "se o aumento e diminución da B. betularia é un dos exemplos máis impactantes visualmente e doadamente comprensibles de evolución darwinista en acción, debería ser ensinado. Proporciona despois de todo a proba da evolución."[49]
Majerus morreu antes de que puidese completar a redacción do resultado dos seus experimentos, polo que ese labor final foi levado a cabo por Cook, Grant, Saccheri e Mallet, e foron publicados o 8 de febreiro de 2012 co título "Selective bird predation on the peppered moth: the last experiment of Michael Majerus." (Predación selectiva polas aves: o último experimento de Michael Majerus)[50] O experimente converteuse no máis grande nunca feito sobre o estudo do melanismo industrial, no cal foron estudados 4864 individuos nunha investigación que durou seis anos, e confirma que o melanismo nas avelaíñas é un exemplo xenuíno de selección natural que implica a camuflaxe e a predación. A súa concluinte observación di: "Estes datos proporcionan a proba máis directa ata agora que implica a camuflaxe e a predación das aves como a explicación principal do aumento e diminución do melanismo en avelaíñas."[8][51]
Coyne respondeu felicitándose de que as súas críticas tivesen motivado a repetición do experimento e dicindo: "A pesar da defensa dos evolucionistas británicos, creo que as miñas críticas tiñan algún peso, porque o biólogo de Cambridge Michael Majerus decidiu repetir os experimentos de Kettlewell, pero facéndoos correctamente esta vez." Citou a conclusión de Cook et al. de que "estes novos datos contestan ás críticas aos traballos iniciais e validan a metodoloxía empregada en moitos experimentos de predación previos que utilizaban os troncos de árbores como sitios de descanso. Os novos datos, acoplados co peso de datos que existían previamente mostran convincentemente que 'o melanismo industrial na B. betularia é aínda un dos exemplos máis claros e doadamente comprensibles da evolución darwinista en acción'." Coyne dixo que estaba "encantado de concordar con esta conclusión, que responde ás miñas críticas previas sobre a historia da Biston."[37]
Notas
[editar | editar a fonte]- ↑ 1,0 1,1 Rudge, David W. (2005). "The Beauty of Kettlewell's Classic Experimental Demonstration of Natural Selection". BioScience 55 (4): 369–375. doi:10.1641/0006-3568(2005)055[0369:TBOKCE]2.0.CO;2.
- ↑ 2,0 2,1 2,2 Majerus, Michael E. N. (2008). "Industrial Melanism in the Peppered Moth, Biston betularia: An Excellent Teaching Example of Darwinian Evolution in Action" (PDF). Evolution: Education and Outreach 2 (1): 63–74. doi:10.1007/s12052-008-0107-y. Arquivado dende o orixinal (PDF) o 08 de agosto de 2017. Consultado o 02 de marzo de 2020.
- ↑ 3,0 3,1 Rice, Stanley A. (2007). Encyclopedia of Evolution. New York: Facts On File. p. 308. ISBN 978-1-4381-1005-9.
- ↑ M. E. N. Majerus (1998). Melanism: Evolution in Action. Oxford University Press. ISBN 978-0-19-854983-3.
- ↑ Clarke, C. A.; Mani, G. S.; Wynne, G. (1985). "Evolution in reverse: clean air and the peppered moth". Biological Journal of the Linnean Society 26 (2): 189–199. doi:10.1111/j.1095-8312.1985.tb01555.x.
- ↑ Majerus, Michael E. N. (2008). "Industrial Melanism in the Peppered Moth, Biston betularia: An Excellent Teaching Example of Darwinian Evolution in Action". Evolution: Education and Outreach 2 (1): 63–74. doi:10.1007/s12052-008-0107-y.
- ↑ 7,0 7,1 7,2 Miller, Ken (1999). The Peppered Moth: An Update
- ↑ 8,0 8,1 Cook, L. M.; Grant, B. S.; Saccheri, I. J.; Mallet, James (2012). "Selective bird predation on the peppered moth: the last experiment of Michael Majerus". Biology Letters 8 (4): 609–612. PMC 3391436. PMID 22319093. doi:10.1098/rsbl.2011.1136.
- ↑ Berry, R. J. (1990). "Industrial melanism and peppered moths (Biston betularia (L.))". Biological Journal of the Linnean Society 39 (4): 301–322. doi:10.1111/j.1095-8312.1990.tb00518.x.
- ↑ Saccheri, I. J.; Rousset, F.; Watts, P. C.; Brakefield, P. M.; Cook, L. M. (2008). "Selection and gene flow on a diminishing cline of melanic peppered moths". Proceedings of the National Academy of Sciences 105 (42): 16212–16217. Bibcode:2008PNAS..10516212S. PMC 2571026. PMID 18854412. doi:10.1073/pnas.0803785105.
- ↑ Neal, Dick (2004). Introduction to Population Biology (Reprint ed.). Cambridge: Cambridge University Press. p. 171. ISBN 9780521532235.
- ↑ Edleston, R. S. (1864). "[No title]". Entomologist 2: 150.
- ↑ 13,0 13,1 Hart, Adam G.; Stafford, Richard; Smith, Angela L.; Goodenough, Anne E. (2010). "Evidence for contemporary evolution during Darwin's lifetime". Current Biology 20 (3): R95. PMID 20144776. doi:10.1016/j.cub.2009.12.010.
- ↑ Farn, A. B. (18 November 1878). "Farn, A.B. to Darwin C.R. , Darwin Correspondence Project Letter 11747". The Darwin Papers. Manuscripts Room, Cambridge University Library, West Road, Cambridge, England. DAR 164:26.
- ↑ "How an extraordinary letter to Darwin spotted industrial melanism in moths". Science Focus. 27 February 2018. Consultado o 16 June 2018.
- ↑ Steward, R. C. (1977). "Industrial and non-industrial melanism in the peppered moth Biston betularia (L.)". Ecological Entomology 2 (3): 231–243. doi:10.1111/j.1365-2311.1977.tb00886.x.
- ↑ http://www.talkorigins.org/faqs/wells/images/grantfile.jpg
- ↑ 18,0 18,1 Cook, L. M. (2003). "The rise and fall of the Carbonaria form of the peppered moth". The Quarterly Review of Biology 78 (4): 399–417. PMID 14737825. doi:10.1086/378925.
- ↑ "University of Chicago Press Journals". PMID 14737825. doi:10.1086/378925.
- ↑ 20,0 20,1 20,2 20,3 Majerus, Michael E. N. (August 2007). "The Peppered Moth: The Proof of Darwinian Evolution" (PDF). Arquivado dende o orixinal (PDF) o 15 de xuño de 2011. Consultado o 2007-09-09.
- ↑ Haldane, J.B.S. (1924). A Mathematical Theory of Natural and Artificial Selection.
- ↑ Bowler, Peter J. (2003). Evolution: The history of an idea, 3rd edition. University of California Press. pp. 331–332. ISBN 0-520-23693-9.
- ↑ Grant, B. S. (2004). "Allelic melanism in American and British peppered moths". Journal of Heredity 95 (2): 97–102. PMID 15073224. doi:10.1093/jhered/esh022.
- ↑ van't Hof, Arjen E.; Edmonds, Nicola; Dalikova, Martina; Marec, Frantisek; Saccheri, Ilik J. (20 May 2011). "Industrial Melanism in British Peppered Moths Has a Singular and Recent Mutational Origin". Science 332 (6032): 958–960. Bibcode:2011Sci...332..958V. PMID 21493823. doi:10.1126/science.1203043 – vía JSTOR.
- ↑ 25,0 25,1 25,2 25,3 25,4 van't Hof, Arjen E.; Campagne, Pascal; Rigden, Daniel J.; Yung, Carl J.; Lingley, Jessica; Quail, Michael A.; Hall, Neil; Darby, Alistair; Saccheri, Ilik J. (2 June 2016). "The industrial melanism mutation in British peppered moths is a transposable element". Nature 534 (7605): 102–117. Bibcode:2016Natur.534..102H. PMID 27251284. doi:10.1038/nature17951.
- ↑ Dobzhansky, T.G. (1937). Genetics and the Evolutionary Process. Columbia University Press. ISBN 0-231-08306-8.
- ↑ Riley, P.A. (2013). "A proposed selective mechanism based on metal chelation in industrial melanic moths" (PDF). Biological Journal of the Linnean Society 109 (2): 298–301. doi:10.1111/bij.12062.
- ↑ A. D. Peacock . John William Heslop Harrison. 1881-1967. Biographical Memoirs of Fellows of the Royal Society. Royal Society. Vol. 14 (Nov., 1968), pp. 243-270 [1]
- ↑ Karl Cleve. Die Erforschung der Ursachen für das Auftreten melanistischer Schmetterlingformen im Laufe der letzten hundert Jahre. Journal of Applied Entomology. 65(1‐4):371 - 387 · August 2009. DOI: 10.1111/j.1439-0418.1970.tb03998.x [2]
- ↑ E. B. Ford. Ecological Genetics
- ↑ Fisher, R. A. (1933). "On the Evidence Against the Chemical Induction of Melanism in Lepidoptera". Proceedings of the Royal Society B 112 (778): 407–416. Bibcode:1933RSPSB.112..407F. doi:10.1098/rspb.1933.0018.
- ↑ Kettlewell, H B D (1958). "A survey of the frequencies of Biston betularia (L.) (Lep.) and its melanic forms in Great Britain". Heredity 12 (1): 51–72. doi:10.1038/hdy.1958.4.
- ↑ Sargent, T. D. (1968). "Cryptic moths: effects on background selections of painting the circumocular scales". Science 159 (3810): 100–101. Bibcode:1968Sci...159..100S. PMID 5634373. doi:10.1126/science.159.3810.100.
- ↑ Sargent, T. D. (1969). "Background Selections of the Pale and Melanic Forms of the Cryptic Moth, Phigalia titea (Cramer)". Nature 222 (5193): 585–586. Bibcode:1969Natur.222..585S. doi:10.1038/222585b0.
- ↑ Sargent, T.D.; Millar, C.D.; Lambert, D.M. (1988). "Ch. 9: The 'classical' explanation of industrial melanism: Assessing the evidence". En Hecht, Max K.; Wallace, Bruce. Evolutionary Biology 23. Plenum Press. ISBN 0306429772.
- ↑ Coyne, Jerry A. (1998). "Not Black and White. Review of Melanism: Evolution in Action by Michael E.N. Majerus". Nature 396 (6706): 35–36. doi:10.1038/23856.
for the time being we must discard Biston as a well-understood example of natural selection in action, although it is clearly a case of evolution. There are many studies more appropriate for use in the classroom.
- ↑ 37,0 37,1 Coyne, Jerry (12 February 2012). "The peppered moth story is solid". Why Evolution Is True. Consultado o 7 March 2012.
Despite the defensiveness of British evolutionists, I think my criticisms carried some weight, because Cambridge biologist Michael Majerus decided to repeat Kettlewell's experiments, but doing them correctly this time." "These new data answer criticisms of earlier work and validate the methodology employed in many previous predation experiments that used tree trunks as resting sites. The new data, coupled with the weight of previously existing data convincingly show that 'industrial melanism in the peppered moth is still one of the clearest and most easily understood examples of Darwinian evolution in action'." [I was] "delighted to agree with this conclusion, which answers my previous criticisms about the Biston story.
- ↑ Kenney, Michael (22 October 2002). "Of Dark Moths, Men and Evolution". Chicago Tribune. Arquivado dende o orixinal o 10 de decembro de 2014. Consultado o 10 December 2014.
- ↑ "Of Moths and Men". W. W. Norton & Company. Arquivado dende o orixinal o 10 de decembro de 2014. Consultado o 10 December 2014.
- ↑ Smith, Peter D. (11 May 2002). "Of Moths and Men: Intrigue, Tragedy & the Peppered Moth". The Guardian. Consultado o 10 December 2014.
- ↑ "Of moths and men". The Independent. 4 September 2003. Consultado o 10 December 2014.
- ↑ Dover, Gabby (2003). "Mothbusters". EMBO Reports 4 (3): 235. PMC 1315906. doi:10.1038/sj.embor.embor778.
- ↑ Grant, B. S. (2002). "Sour grapes of wrath". Science 297: 940–941. doi:10.1126/science.1073593.
- ↑ Majerus, Michael E. N. (2005). "The peppered moth: decline of a Darwinian disciple". En Fellowes, Mark; Holloway, Graham; Rolf, Jens. Insect Evolutionary Ecology. Wallingford, Oxon: CABI Publishing. pp. 375–377. ISBN 978-1-84593-140-7.
- ↑ Frack, Donald (16 April 1999). "Evolution – April 1999: Peppered Moths and Creationists". Arquivado dende o orixinal o 26 de agosto de 2007. Consultado o 2007-08-26.
- ↑ Wells, J. (24 May 1999). "Second Thoughts about Peppered Moths; This classical story of evolution by natural selection needs revising". The Scientist 13 (11): 13.
- ↑ Wells J. (2000). Icons of Evolution: Science or Myth? Why Much of What We Teach About Evolution is Wrong. Regnery Press, Washington, D.C., p. 138 (book available from Iconsofevolution.com). Orixinal: "What the textbooks don't explain, however, is that biologists have known since the 1980s that the classical story has some serious flaws. The most serious is that peppered moths in the wild don't even rest on tree trunks. The textbook photographs, it turns out, have been staged."
- ↑ "Icon of Obfuscation: Chapter 7: Peppered Moths".
- ↑ Connor, Steve (25 August 2007). "Moth study backs classic 'test case' for Darwin's theory". The Independent. Arquivado dende o orixinal o 07 de outubro de 2008. Consultado o 2007-09-09.
- ↑ Matzke, Nick (8 February 2012). "Selective bird predation on the peppered moth: the last experiment of Michael Majerus". The Panda's Thumb. Arquivado dende o orixinal o 29 de setembro de 2019. Consultado o 7 March 2012.
- ↑ Orixinal en Cook2012: "These data provide the most direct evidence yet to implicate camouflage and bird predation as the overriding explanation for the rise and fall of melanism in moths."
{kind=link}
Véxase tamén
[editar | editar a fonte]| Commons ten máis contidos multimedia sobre: Evolución de Biston betularia |
Outros artigos
[editar | editar a fonte]Ligazóns externas
[editar | editar a fonte]- Bruce Grant escribiu varios artigos sobre melanismo en B. betularia que están listados na súa páxina web.
- Conferencia en liña: "The rise and fall of the melanic Peppered Moth" de Laurence Cook.
- Young, Matt. "Moonshine: Why the Peppered Moth remains an Icon of Evolution". Department of Physics, Colorado School of Mines. Consultado o 2009-01-03.
- The Peppered Moth: Decline of a Darwinian Disciple. Este é o transcrito da conferencia de Michael Majerus que deu no British Humanist Association no día de Darwin de 2004.
- The Peppered Moth: The Proof of Darwinian Evolution. Este é a transcrición da conferencia de Majerus dada na xuntanza European Society for Evolutionary Biology o 23 de agusto de 2007. Tamén inclúe unha presentación en powerpoint [3].
- Majerus, Michael E. N. (2009). "Industrial Melanism in the Peppered Moth, Biston betularia: An Excellent Teaching Example of Darwinian Evolution in Action". Evolution: Education and Outreach 2 (1): 63–74. doi:10.1007/s12052-008-0107-y.
As acusacións de falseamento de datos e fraude científica no caso reveláronse sen fundamento.
- O 19 de xuño de 2009, Telegraph.co.ukArquivado 25 de decembro de 2019 en Wayback Machine. publicou este artigoArquivado 01 de novembro de 2013 en Wayback Machine. sobre este fenómeno evolutivo e invitaba aos lectores británicos a visitar a páxina web Moths Count e rexistrar as súas observacións de avelaíñas locais, para axudar aos investigadores a incrementar os datos dispoñibles.